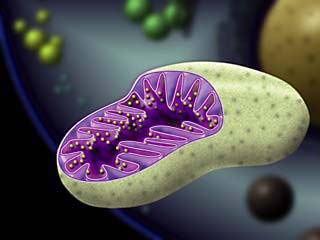
MİTOKONDRİ
Eukaryotik
bütün hayvansal ve bitkisel hücrelerin sitoplazmasında bulunan, granüler veya
ipliksi, küçük organellere mitokondriyum denir. Normal bir karaciğer hücresinde
1000-1600, bazı oositlerde 300.000 kadar mitokondri bulunur. Yeşil bitkiler
hayvansal hücrelere kıyasla çok az mitokondri içerirler. Çünkü mitokondrilerin
bazı görevleri kloroplastlar tarafından yerine getirilir.
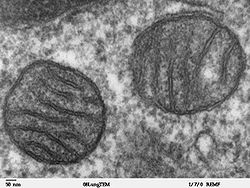
MİTOKONDRİLERİN BULUNUŞU
Mitokondriler ilk olarak KOLLİCKER tarafından böbrek çizgili kas hücrelerinden izole edilmişler ve değişik tuzlu solusyonlar içinde bu yapıların osmotik reaksiyonları yaklaşık 150 yıl önce, o dönemlerde gözlenebilmiştir. 1850’den itibaren yayınlanan çalışmalarla beraber bu organelin ilk tanımlanması, 1894’de ALTMANN tarafından yapılmıştır. Işık mikroskobu ile hücrede granüller veya iplikler şeklinde görülen bu organele 1897’de BENDA, mitokondrion adını vermiştir. (Y.mitos: iplik ; chondrion :granül).
Mitokondriler canlı olarak ilk defa 1900 yılında MICHAELES’in kullandığı, bugün de kullanılan ve vital boya olan Janus yeşili B ile mavimsi yeşile boyanarak diğer hücre organellerinden kolaylıkla ayırt edilirler. Janus yeşili mitokondriler için özeldir. Bu boya için mitokondrilerin yeşile boyanması mitokondri içinde bulunan sitokrom oksidaz sisteminin boyayı oksitlenmiş halde renkli tutması, çevredeki sitoplazmada ise boyanın renksiz olan lökobaza redüklenmesinden ileri gelmektedir.
1913’de, O. WARBURG solunum enzimlerinin mitokondrilerde olduğunu gözlemiştir. Mitokondrileri düşük devirde santrifüj ederek dokulardan izole eden WARBURG, dokuların ezilerek parçalanması sonucunda bu organellerin hücre içi oksidatif reaksiyonları katalizleyen enzimler içerdiğini görmüştür. WARBURG ve WIELAND, 1920’lerde sitokrom oksidaz olayını ve Krebs Devri olarak bilinen ve küçük karbon zincirlerinin parçalanarak devamlı bir şekilde CO2 in meydana geldiği reaksiyonlar devrini açıklamıştır. KALCKAR, 1940’da solunum zinciri reaksiyonlarında oksijenin suya indirgendiğini ve yeni ATPler teşekkül ettiğini açıklamış, buna oksidatif fosforilasyon demiştir.
1934’de, BENSLEY ve HOERR, karaciğerden mitokondrileri ayırmışlar, organelin biyokimyasal olarak incelenme imkanı ortaya çıkmıştır. Canlı hücreler, ultrasantrifüjde 200.000-400.000 devirle santrifüje edilirlerse, santrifüj tüpünün dibinde, sitoplazmadan daha ağır ve yoğun olan mitokondrilerin parçalanmamış olarak toplanması sağlanır. Kimyasal analizler sonucu, bileşiminde %65-70 protein, %25-30 lipit, %2-3 kolesterol olduğu gösterilmiştir. Ayrıca solunum enzimleri, nükleik asitler (DNA ve RNA) nükleotitlerin (ATP) bulunduğu da öğrenilmiştir.
G. H. HOGEBOOM ve W. C. SCHNEIDER, 1948’de sitokrom faaliyetinin çoğunun gerçekten bu hücre fraksiyonlarında meydana geldiğini göstermişlerdir.
1950’li yıllarda bu reaksiyonlar A. L. LEHNINGER, G. H. HOGEBOOM tarafından trikarboksilik asit reaksiyonları gözlenmiş, hem yağ asitlerinin oksidasyonunun hem de oksidatif fosforilasyonun mitokondrilerde meydana geldiği kesin olarak açıklanmıştır.
1952’de PALADE ve SJÖSTRAND, elektron mikroskobu ile organelin ince yapısı üzerinde ilk bilgileri sağlamışlardır. Son yıllarda geliştirilen birçok teknikle organel çok daha iyi öğrenilmiştir.
Mitokondriler canlı hücrelerde incelenebilirler, ancak alçak kırılma indisleri nedeniyle boyanmadıkça kolay görünemezler. Doku kültürü hücrelerinde en iyi biçimde faz kontrast mikroskobu ile incelenebilirler.
Şekil 1.Mısır (Zea mays) kök ucunda meristem hücrelerindeki mitokondriyumlar. N, nukleus; nu, nukleolus; mi, mitokondriyumlar.
Mitokondriler canlı olarak ilk defa 1900 yılında MICHAELES’in kullandığı, bugün de kullanılan ve vital boya olan Janus yeşili B ile mavimsi yeşile boyanarak diğer hücre organellerinden kolaylıkla ayırt edilirler. Janus yeşili mitokondriler için özeldir. Bu boya için mitokondrilerin yeşile boyanması mitokondri içinde bulunan sitokrom oksidaz sisteminin boyayı oksitlenmiş halde renkli tutması, çevredeki sitoplazmada ise boyanın renksiz olan lökobaza redüklenmesinden ileri gelmektedir.
1913’de, O. WARBURG solunum enzimlerinin mitokondrilerde olduğunu gözlemiştir. Mitokondrileri düşük devirde santrifüj ederek dokulardan izole eden WARBURG, dokuların ezilerek parçalanması sonucunda bu organellerin hücre içi oksidatif reaksiyonları katalizleyen enzimler içerdiğini görmüştür. WARBURG ve WIELAND, 1920’lerde sitokrom oksidaz olayını ve Krebs Devri olarak bilinen ve küçük karbon zincirlerinin parçalanarak devamlı bir şekilde CO2 in meydana geldiği reaksiyonlar devrini açıklamıştır. KALCKAR, 1940’da solunum zinciri reaksiyonlarında oksijenin suya indirgendiğini ve yeni ATPler teşekkül ettiğini açıklamış, buna oksidatif fosforilasyon demiştir.
1934’de, BENSLEY ve HOERR, karaciğerden mitokondrileri ayırmışlar, organelin biyokimyasal olarak incelenme imkanı ortaya çıkmıştır. Canlı hücreler, ultrasantrifüjde 200.000-400.000 devirle santrifüje edilirlerse, santrifüj tüpünün dibinde, sitoplazmadan daha ağır ve yoğun olan mitokondrilerin parçalanmamış olarak toplanması sağlanır. Kimyasal analizler sonucu, bileşiminde %65-70 protein, %25-30 lipit, %2-3 kolesterol olduğu gösterilmiştir. Ayrıca solunum enzimleri, nükleik asitler (DNA ve RNA) nükleotitlerin (ATP) bulunduğu da öğrenilmiştir.
G. H. HOGEBOOM ve W. C. SCHNEIDER, 1948’de sitokrom faaliyetinin çoğunun gerçekten bu hücre fraksiyonlarında meydana geldiğini göstermişlerdir.
1950’li yıllarda bu reaksiyonlar A. L. LEHNINGER, G. H. HOGEBOOM tarafından trikarboksilik asit reaksiyonları gözlenmiş, hem yağ asitlerinin oksidasyonunun hem de oksidatif fosforilasyonun mitokondrilerde meydana geldiği kesin olarak açıklanmıştır.
1952’de PALADE ve SJÖSTRAND, elektron mikroskobu ile organelin ince yapısı üzerinde ilk bilgileri sağlamışlardır. Son yıllarda geliştirilen birçok teknikle organel çok daha iyi öğrenilmiştir.
Mitokondriler canlı hücrelerde incelenebilirler, ancak alçak kırılma indisleri nedeniyle boyanmadıkça kolay görünemezler. Doku kültürü hücrelerinde en iyi biçimde faz kontrast mikroskobu ile incelenebilirler.
Şekil 1.Mısır (Zea mays) kök ucunda meristem hücrelerindeki mitokondriyumlar. N, nukleus; nu, nukleolus; mi, mitokondriyumlar.
MİTOKONDRİLERİN KÖKENİ
Hücrede mitokondrilerin teşekkülü ile ilgili olarak üç görüş ortaya atılmıştır. Buna göre:
1) Mevcut mitokondrilerin bölünmesi ile
2) Yeniden sentez edilmeleri ile
3) Veya başka yapıların gelişmesiyle” oluşmuşlardır.
Eski bir görüşe göre mitokondrilerin kromozomlar gibi kalıtımda rol oynadıkları, kromozomlar gibi bölünüp çoğaldıktan sonra yavru hücrelere geçtikleri kabul ediliyordu. Gerçekten de doku kültürlerinde mitokondrilerin birleştikleri gibi, yavaş yavaş büyüdükten sonra parçalandıkları görülmüş, mitozda yavru hücrelere bölünerek interfazda sayılarını arttırdıkları gözlenmiştir. Bu gözlem kolin eksikliği olan Neurospora’da yapılan bir deneyle doğrulanmıştır. Neurospora’ya radyoaktif kolin verildiği takdirde orijinal mitokondrilerden gelişen 2. ve 3. jenerasyondaki mitokondrilerde otoradyografi ile radyoaktivite saptanmıştır. Mitokondrilerin radyoaktivitesi her dölde yarı yarıya azalma göstermektedir. Bu, mitokondrilerin bölünerek çoğaldığını ve mitokondrilerin temel yapısına yeni maddelerin ilave edildiğini göstermektedir.
Elektron mikroskobu çalışmalarından sonra mitokondrilerin plazma zarından meydana geldiği kabul edilmiştir. Bira mayasının anaerobik kültüründe mitokondrilere rastlanmadığı halde, bu hücreler aerobik koşulara maruz bırakılacak olursa plazma zarlarının birleşip katlanarak gerçek mitokondrileri meydana getirdiği görülür. Bu gözlem mitokondrilerin yeniden sentez edildiğini savunan teoriyle bir uygunluk göstermektedir. Mitokondri dış zarının kimyasal yapısı ER zarına benzer ve iç zardan farklıdır. Dış zar lipid sentezinde iç zardan daha aktiftir. Mitokondrilerin dış zarı ile ER’un ilişkisi de elektron mikroskobu ile gözlenmiştir. Biyokimyasal deliller ile karaciğer mitokodrilerinin yarı ömürlerinin 5-10 gün olduğu gösterilmiştir. Bu gözlem hücrede mitokondri oluşumunun devamlı olarak meydana geldiğini ortaya koymaktadır.
Mitokondriler ancak birkaç gün yaşar. Enine bölünme ile çoğalır. Bunlar embriyonel hücrelerdeki promitokondrilerden de gelişir ve çok küçük vesiküller olup yoğun matriksli, çift membranlıdır. İç membranlarında zamanla kristalar gelişir. Promitokondriler, bölünme ile kendilerinden ve tomurcuklanma ile gelişmiş mitokondrilerden oluşur. Mitokondrinin ortalama ömrü hücreye göre daha azdır. Karaciğer hücre mitokondrileri 7-10 gün yaşar.
Sinematografi kullanılarak mitokondrilerin bölünme işlemlerinin incelenmesi ile bölünmeye başlarken mitokondrilerin uzadıkları, merkeze yakın bir bölgeden haltere benzer bir görünüm alacak şekilde boğumlandıkları, bu bölgeden ikiye bölündükleri, bölünmeden sonra da yeniden yuvarlak şekil aldıkları görülmektedir.
L.MARGULİS eukaryotik hücrelerdeki mitokondrilerin evrimsel süreçte prokaryotik organizmalardan geliştiklerine dair açıklamalarda bulunmuştur. Bu düşüncesini mitokondrilerde varolan DNA ve mitoribozomlara dayandırmıştır. Mitokondriler sahip oldukları DNA ve ribozomlar sayesinde hücrede bağımsız olarak nükleer ve sitoplazmik ribozomal işlemlere ait fonksiyonları yerine getirirler. MARGULİS’in bu fikri mitokondrial DNA’nın prokaryotik DNA’ya benzemesi, prokaryotik DNA gibi tek zincirli molekül olması mitokondrial ribozomların; sedimentasyon katsayılarından anlaşıldığı gibi protein ve RNA bileşimlerinden meydana gelmeleri ve prokaryotik ribozomlara ökaryotik ribozomlardan daha fazla benzemeleri gibi durumlarla daha fazla desteklenmektedir.
1) Mevcut mitokondrilerin bölünmesi ile
2) Yeniden sentez edilmeleri ile
3) Veya başka yapıların gelişmesiyle” oluşmuşlardır.
Eski bir görüşe göre mitokondrilerin kromozomlar gibi kalıtımda rol oynadıkları, kromozomlar gibi bölünüp çoğaldıktan sonra yavru hücrelere geçtikleri kabul ediliyordu. Gerçekten de doku kültürlerinde mitokondrilerin birleştikleri gibi, yavaş yavaş büyüdükten sonra parçalandıkları görülmüş, mitozda yavru hücrelere bölünerek interfazda sayılarını arttırdıkları gözlenmiştir. Bu gözlem kolin eksikliği olan Neurospora’da yapılan bir deneyle doğrulanmıştır. Neurospora’ya radyoaktif kolin verildiği takdirde orijinal mitokondrilerden gelişen 2. ve 3. jenerasyondaki mitokondrilerde otoradyografi ile radyoaktivite saptanmıştır. Mitokondrilerin radyoaktivitesi her dölde yarı yarıya azalma göstermektedir. Bu, mitokondrilerin bölünerek çoğaldığını ve mitokondrilerin temel yapısına yeni maddelerin ilave edildiğini göstermektedir.
Elektron mikroskobu çalışmalarından sonra mitokondrilerin plazma zarından meydana geldiği kabul edilmiştir. Bira mayasının anaerobik kültüründe mitokondrilere rastlanmadığı halde, bu hücreler aerobik koşulara maruz bırakılacak olursa plazma zarlarının birleşip katlanarak gerçek mitokondrileri meydana getirdiği görülür. Bu gözlem mitokondrilerin yeniden sentez edildiğini savunan teoriyle bir uygunluk göstermektedir. Mitokondri dış zarının kimyasal yapısı ER zarına benzer ve iç zardan farklıdır. Dış zar lipid sentezinde iç zardan daha aktiftir. Mitokondrilerin dış zarı ile ER’un ilişkisi de elektron mikroskobu ile gözlenmiştir. Biyokimyasal deliller ile karaciğer mitokodrilerinin yarı ömürlerinin 5-10 gün olduğu gösterilmiştir. Bu gözlem hücrede mitokondri oluşumunun devamlı olarak meydana geldiğini ortaya koymaktadır.
Mitokondriler ancak birkaç gün yaşar. Enine bölünme ile çoğalır. Bunlar embriyonel hücrelerdeki promitokondrilerden de gelişir ve çok küçük vesiküller olup yoğun matriksli, çift membranlıdır. İç membranlarında zamanla kristalar gelişir. Promitokondriler, bölünme ile kendilerinden ve tomurcuklanma ile gelişmiş mitokondrilerden oluşur. Mitokondrinin ortalama ömrü hücreye göre daha azdır. Karaciğer hücre mitokondrileri 7-10 gün yaşar.
Sinematografi kullanılarak mitokondrilerin bölünme işlemlerinin incelenmesi ile bölünmeye başlarken mitokondrilerin uzadıkları, merkeze yakın bir bölgeden haltere benzer bir görünüm alacak şekilde boğumlandıkları, bu bölgeden ikiye bölündükleri, bölünmeden sonra da yeniden yuvarlak şekil aldıkları görülmektedir.
L.MARGULİS eukaryotik hücrelerdeki mitokondrilerin evrimsel süreçte prokaryotik organizmalardan geliştiklerine dair açıklamalarda bulunmuştur. Bu düşüncesini mitokondrilerde varolan DNA ve mitoribozomlara dayandırmıştır. Mitokondriler sahip oldukları DNA ve ribozomlar sayesinde hücrede bağımsız olarak nükleer ve sitoplazmik ribozomal işlemlere ait fonksiyonları yerine getirirler. MARGULİS’in bu fikri mitokondrial DNA’nın prokaryotik DNA’ya benzemesi, prokaryotik DNA gibi tek zincirli molekül olması mitokondrial ribozomların; sedimentasyon katsayılarından anlaşıldığı gibi protein ve RNA bileşimlerinden meydana gelmeleri ve prokaryotik ribozomlara ökaryotik ribozomlardan daha fazla benzemeleri gibi durumlarla daha fazla desteklenmektedir.
MİTOKONDRİLERİN BİÇİM ve BÜYÜKLÜKLERİ
Mitokondrilerin yapısı, biçimi, büyüklüğü, sayısı ve hücredeki yerleri, bulunduğu dokunun fizyolojik durumuna bağlı olarak hücreden hücreye değişiklik gösterir. Genellikle küresel, ovoid, silindirik, çomak ya da granül biçiminde olurlar. Bununla birlikte çomak biçimindeki mitokondrilerin parçalanarak granül, granül biçimindeki mitokondrilerin birbirleriyle birleşerek çomak biçimindeki mitokondrileri yaptıkları da gözlenmektedir.
Mitokondrilerin büyüklükleri de değişik olur. Pek çok hücrede çapları oldukça sabit(ortalama 0.5 µm), fakat uzunlukları farklıdır. Çomak biçimindeki mitokondriler 0.5µm-1µm çapında ve 2-3µm uzunluğunda olurlar. Az da olsa 7-9µm uzunluğunda mitokondrilere de rastlanır.
Mitokondrilerin biçim ve büyüklükleri kullanılan fiksatifin PH'ına ve osmatik basıncına bağlıdır. Genellikle sitoplazmaya homojen olarak yayılmalarına rağmen nukleus etrafında veya çevresel sitoplazmada toplanabilirler. Sitoplazmada mitokondri dağılımının, oksidasyon merkezi olarak kabul edilen bu organellerin gerekli enerji sağlamak için ATP'ı hücrenin gerek duyulan bölgesine iletmesiyle ilgili olduğu düşünülmektedir. Örneğin kas hücrelerinde mitokondriler halka halinde I- bandının etrafında yer alırlar. Sitoplazma hareketine bağlı olarak yerlerini pasif şekilde değiştirirler. Hücre bölünmesinde iğe yakın bölgelerde toplanırlar ve hücre bölünürken yavru hücrelere aşağı yukarı eş miktarda dağılırlar.
Şekil 2. Mitokondri ince yapı değişiklikleri.
Mitokondrilerin büyüklükleri de değişik olur. Pek çok hücrede çapları oldukça sabit(ortalama 0.5 µm), fakat uzunlukları farklıdır. Çomak biçimindeki mitokondriler 0.5µm-1µm çapında ve 2-3µm uzunluğunda olurlar. Az da olsa 7-9µm uzunluğunda mitokondrilere de rastlanır.
Mitokondrilerin biçim ve büyüklükleri kullanılan fiksatifin PH'ına ve osmatik basıncına bağlıdır. Genellikle sitoplazmaya homojen olarak yayılmalarına rağmen nukleus etrafında veya çevresel sitoplazmada toplanabilirler. Sitoplazmada mitokondri dağılımının, oksidasyon merkezi olarak kabul edilen bu organellerin gerekli enerji sağlamak için ATP'ı hücrenin gerek duyulan bölgesine iletmesiyle ilgili olduğu düşünülmektedir. Örneğin kas hücrelerinde mitokondriler halka halinde I- bandının etrafında yer alırlar. Sitoplazma hareketine bağlı olarak yerlerini pasif şekilde değiştirirler. Hücre bölünmesinde iğe yakın bölgelerde toplanırlar ve hücre bölünürken yavru hücrelere aşağı yukarı eş miktarda dağılırlar.
Şekil 2. Mitokondri ince yapı değişiklikleri.
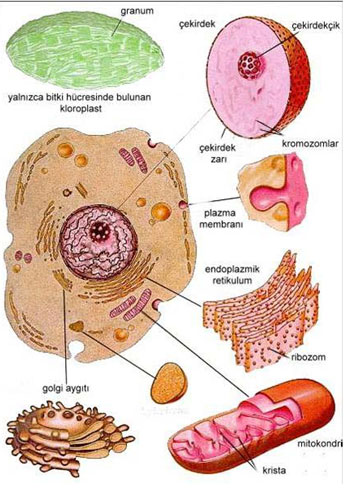
MİTOKONDRİLERİN SAYISI
Mitokondrilerin sayısı hücrenin ve organelin aktivitesine bağlıdır. Bakteri ve mavi-yeşil algler gibi prokaryotik organizmalarda mitokondri bulunmadığı halde sitosolde onun görevini sitosolde mitokondri benzeri yapılar yada plazma membranı yerine getirmektedir. Bir fungus olan Blastocladiella emersonia da ve bir alg olan Chlorella jusca da tek bir mitokondri bulunmaktadır. Amip olan Choas'ta ise yüzlerce mitokondri vardır.
Yüksek hayvansal organizmaların hücrelerinde genelde farklı sayıda mitokondri bulunmaktadır. Örneğin sperm hücrelerinde yaklaşık 100 kadar, böbrek hücrelerinde 1000 kadar mitokondri bulunur.
İnce barsak yüzeyini astarlayan epitel hücrelerinin hücre yüzeyine yakın bölgelerinde çok fazla sayıda bulunan mitokondriler absorbsiyonda gerekli enerjiyi sağlamaktadırlar.
Hücrelerdeki mitokondri sayısının fazla olması o hücrelerde ATP kullanımının çok olduğunu göstermektedir.
İNCE YAPISI ve KRİSTALAR
Mitokondrilerin ince yapısını elektron mikroskobuyla görmek mümkündür. Işık mikroskobunda incelenmeleri için ozmik asit, potasyum bikromat kullanılır. Fuksin asiti ve demir hematoksilin de genellikle kullanılan boyalardır. Organel, dış taraftan her biri 60-80 Aº kalınlığında ve 60-80 Aº aralıkla birbirine paralel iki zarla çevrilidir. Her iki zar da plazma zarı gibi " birim zar" özelliği gösterir, protein, lipid ,protein katlarından yapılıdır. Bununla birlikte plazma zarı ile mitokondri dış ve iç zarları arasında yapı ve fonksiyon bakımından farklar vardır.
PALADE ve SJÖSTRAND'ın elektron mikroskobu çalışmalarıyla organelin ince yapısı incelenmiştir. İçteki zarın mitokondrinin iç boşluğuna uzanan ve genellikle yassı keseler şeklinde olan kompleks katlanmalarına krista adı verilmiştir. Memeli hayvan hücresinde yaprak benzeri veya tüpe benzeyen kristalar varken,bitkilerde cep şeklinde sacculi denen yapılar vardır.
Şekil 5. Bir mitokondri kristasının görünümü. 1. Mitokondrinin dış zarı. 2. Mitokondrinin iç zarı. 3. Krista. 4. Küremsi başlıklı partiküller. 5. Başlıkları iç zara bağlayan silindirik boyun kısmı.
Mitokondrilerde dış zar, iç zar, krista ve matriks
İç membran sadece belirli bazı moleküller için geçirgendir. (Mol ağırlığı<0.1 kg/mol). Daha çok transport proteini içerir ve proteince zengindir. (%75 protein, %25 lipid). Dış membran gibi kolesterol içermez. İç membranda ve cristada entegre olarak elektron transportu için enzim kompleksleri bulunur.
Son yapılan araştırmalarda, mantarlarda (BRACKER ve GROVE, 1971, eukaryotik hücrelerde, mitokondrilerin dış zarıyla ER zarlarının bağlantı gösterdiği gözlenmiştir. Bazen mitokondrilere bağlı sistemlerin az miktarda ribozom da içerdiği gözlenmiştirBu sistemlerin özel tipte proteinlerinin ER'den mitokondrilere iletimde rol oynadığı düşünülmüştür.
Karaciğer, böbrek, sinir hücrelerinde krista yapıları.
Dış zarla iç zar ve aynı zamanda kristalar içinde devam eden dış bölmenin zarlar arasındaki kısmına periferal bölge (zarlararası bölge), kristalar içindeki kısmına intrakristal bölge denir. İkinci bölme mitokondri iç zarının çevrelediği kristaların arasında kalan iç bölmedir. Bu kısma mitokondri matriksi denir.
Matriks genelde homojen görünse de bazen çok ince filamentlerden oluşur ve yoğun granüller içerir. Bu granüllerin Ca² ve Mg² iyonlarını bağlayan yerler olduğu düşünülmektedir. Büyük granüller ribozomlardan 2-3 defa büyük olan yuvarlak yapılardır. Granüllerin bir tipi depo kalsiyum iyonlarıdır. Jele benzer sıvı yapıda olan matrikste tuzlar, su, Krebs çemberi (trikarboksilik asit çemberi)nin, sitrat siklüsünün ve yağ asidi oksidasyonun enzimleri bulunur. İç kısımlarda örneğin piruvat, karboksilleşir veya solunumda dekarboksilleşir; burada mitokondrial sentezlerin çoğu olaylanır. Ayrıca matrikste DNA, RNA ve mitokondri ribozomları olan mitoribozomlar bulunur.
Mitokondri ribozomları total olarak 55 S olmak üzere 35 S ve 25 S alt birimlerinden yapılmışlardır. Fibrillerin 30-40 AO çapında olduğu ve çeşitli tekniklerle bunun halkasal bir DNA ipliği olduğu gösterilmiştir. Bazen büyük protein kristalleri, vitellüs granülleri, glikojen taneleri görülmüş olup bunların da depolanabileceği düşünülmüştür.
Mitokondrinin dış zarı ile iç zarı arasında kalan zarlar arası bölgede adenilat kinaz ve nükleosid difosfokinaz enzimleri bulunmuştur. Bu aralıkla mitokondri iç zarının dış zarla yer yer birleştiği bölgelere temas bölgeleri (kontak bölgeleri) adı verilir.
İlk defa FERNANDEZ-MORAN (1962) tarafından geliştirilen negatif boyama yöntemiyle organelin ince yapısında daha ayrıntılı bir yapının varlığı ortaya çıkarılmıştır.Eğer mitokondriler hipotonik bir ortamda şişmeye bırakılır ve sonra fosfotungstata batırılırsa mitokondri iç zarının matrikse bakan yüzünün 8.5 nm çapında ve birer sapla iç zara bağlı partiküllerle kaplı olduğu matrikse bakan yüzünde 10 nm aralıkla düzenli sıralanırlar. Bu yapılara "F1 partikülü, Elementer partikül veya Racker faktörü" adı verilir. Organelde 104-105 kadar elemanter partikül bulunabilir. Bu partiküllerin baş kısımlarının oksidasyon ve fosforilasyonda rolü olan bir ATPaz olduğu bulunmuştur.
Mitokondri fonksiyonu ile ilgili olarak krista sayısı çok değişik olur. Kalp kası, böbrek tübülleri, çizgili kaslarda ve hayvanların uçma kaslarında, oksidatif fosforilasyon olaylarının arttığı durumlarda çok sık dizilmiş kristalara rastlanır.En fazla krista sayısı böceklerin uçma kaslarında bulunur.
Mitokondriler labil yapılı organellerdir. Farklı etmenlerin etkisinde değişikliğe uğrarlar. Zarar gören hücrelerde parçalanarak küçük taneler haline geçerler. İçlerinde yoğun maddeler birikerek daha şişkin hale gelebilirler. Değişiklikler bir sınırı aşınca mitokondriler dejenerasyona uğrarlar. Bundan sonra meydana gelen değişiklikler artık reversibl değildir. Reversibl olmayan değişiklikler üç şekilde gelişir:
1) Ya küçük tanecikler halinde parçalanarak dispersiyon oluştururlar
2) Veya şişerek geniş vakuoller haline gelirler
3) Ya da mitokondrilerin hiyalin granülleri haline dönüşmesiyle şişmiş, geniş, kitlesel birikintiler meydana getirirler.
Böylelikle çoğunlukla hücre ölür. Normal hücrelerde bir tür lisosom (sitolisosom) oluşumuyla mitokondriler dejenerasyona uğrayara k otolize olurlar. Mitokondrilerin bir başka hali de bir araya gelip birleşerek kondriosfer denen daha büyük cisimleri meydana getirmeleridir. Hacim ve şekil değiştirmeleri, kimyasal maddeler ve osmatik basınç değişiklikleriyle sağlanabilir.
Yüksek hayvansal organizmaların hücrelerinde genelde farklı sayıda mitokondri bulunmaktadır. Örneğin sperm hücrelerinde yaklaşık 100 kadar, böbrek hücrelerinde 1000 kadar mitokondri bulunur.
İnce barsak yüzeyini astarlayan epitel hücrelerinin hücre yüzeyine yakın bölgelerinde çok fazla sayıda bulunan mitokondriler absorbsiyonda gerekli enerjiyi sağlamaktadırlar.
Hücrelerdeki mitokondri sayısının fazla olması o hücrelerde ATP kullanımının çok olduğunu göstermektedir.
İNCE YAPISI ve KRİSTALAR
Mitokondrilerin ince yapısını elektron mikroskobuyla görmek mümkündür. Işık mikroskobunda incelenmeleri için ozmik asit, potasyum bikromat kullanılır. Fuksin asiti ve demir hematoksilin de genellikle kullanılan boyalardır. Organel, dış taraftan her biri 60-80 Aº kalınlığında ve 60-80 Aº aralıkla birbirine paralel iki zarla çevrilidir. Her iki zar da plazma zarı gibi " birim zar" özelliği gösterir, protein, lipid ,protein katlarından yapılıdır. Bununla birlikte plazma zarı ile mitokondri dış ve iç zarları arasında yapı ve fonksiyon bakımından farklar vardır.
PALADE ve SJÖSTRAND'ın elektron mikroskobu çalışmalarıyla organelin ince yapısı incelenmiştir. İçteki zarın mitokondrinin iç boşluğuna uzanan ve genellikle yassı keseler şeklinde olan kompleks katlanmalarına krista adı verilmiştir. Memeli hayvan hücresinde yaprak benzeri veya tüpe benzeyen kristalar varken,bitkilerde cep şeklinde sacculi denen yapılar vardır.
Şekil 5. Bir mitokondri kristasının görünümü. 1. Mitokondrinin dış zarı. 2. Mitokondrinin iç zarı. 3. Krista. 4. Küremsi başlıklı partiküller. 5. Başlıkları iç zara bağlayan silindirik boyun kısmı.
Mitokondrilerde dış zar, iç zar, krista ve matriks
İç membran sadece belirli bazı moleküller için geçirgendir. (Mol ağırlığı<0.1 kg/mol). Daha çok transport proteini içerir ve proteince zengindir. (%75 protein, %25 lipid). Dış membran gibi kolesterol içermez. İç membranda ve cristada entegre olarak elektron transportu için enzim kompleksleri bulunur.
Son yapılan araştırmalarda, mantarlarda (BRACKER ve GROVE, 1971, eukaryotik hücrelerde, mitokondrilerin dış zarıyla ER zarlarının bağlantı gösterdiği gözlenmiştir. Bazen mitokondrilere bağlı sistemlerin az miktarda ribozom da içerdiği gözlenmiştirBu sistemlerin özel tipte proteinlerinin ER'den mitokondrilere iletimde rol oynadığı düşünülmüştür.
Karaciğer, böbrek, sinir hücrelerinde krista yapıları.
Dış zarla iç zar ve aynı zamanda kristalar içinde devam eden dış bölmenin zarlar arasındaki kısmına periferal bölge (zarlararası bölge), kristalar içindeki kısmına intrakristal bölge denir. İkinci bölme mitokondri iç zarının çevrelediği kristaların arasında kalan iç bölmedir. Bu kısma mitokondri matriksi denir.
Matriks genelde homojen görünse de bazen çok ince filamentlerden oluşur ve yoğun granüller içerir. Bu granüllerin Ca² ve Mg² iyonlarını bağlayan yerler olduğu düşünülmektedir. Büyük granüller ribozomlardan 2-3 defa büyük olan yuvarlak yapılardır. Granüllerin bir tipi depo kalsiyum iyonlarıdır. Jele benzer sıvı yapıda olan matrikste tuzlar, su, Krebs çemberi (trikarboksilik asit çemberi)nin, sitrat siklüsünün ve yağ asidi oksidasyonun enzimleri bulunur. İç kısımlarda örneğin piruvat, karboksilleşir veya solunumda dekarboksilleşir; burada mitokondrial sentezlerin çoğu olaylanır. Ayrıca matrikste DNA, RNA ve mitokondri ribozomları olan mitoribozomlar bulunur.
Mitokondri ribozomları total olarak 55 S olmak üzere 35 S ve 25 S alt birimlerinden yapılmışlardır. Fibrillerin 30-40 AO çapında olduğu ve çeşitli tekniklerle bunun halkasal bir DNA ipliği olduğu gösterilmiştir. Bazen büyük protein kristalleri, vitellüs granülleri, glikojen taneleri görülmüş olup bunların da depolanabileceği düşünülmüştür.
Mitokondrinin dış zarı ile iç zarı arasında kalan zarlar arası bölgede adenilat kinaz ve nükleosid difosfokinaz enzimleri bulunmuştur. Bu aralıkla mitokondri iç zarının dış zarla yer yer birleştiği bölgelere temas bölgeleri (kontak bölgeleri) adı verilir.
İlk defa FERNANDEZ-MORAN (1962) tarafından geliştirilen negatif boyama yöntemiyle organelin ince yapısında daha ayrıntılı bir yapının varlığı ortaya çıkarılmıştır.Eğer mitokondriler hipotonik bir ortamda şişmeye bırakılır ve sonra fosfotungstata batırılırsa mitokondri iç zarının matrikse bakan yüzünün 8.5 nm çapında ve birer sapla iç zara bağlı partiküllerle kaplı olduğu matrikse bakan yüzünde 10 nm aralıkla düzenli sıralanırlar. Bu yapılara "F1 partikülü, Elementer partikül veya Racker faktörü" adı verilir. Organelde 104-105 kadar elemanter partikül bulunabilir. Bu partiküllerin baş kısımlarının oksidasyon ve fosforilasyonda rolü olan bir ATPaz olduğu bulunmuştur.
Mitokondri fonksiyonu ile ilgili olarak krista sayısı çok değişik olur. Kalp kası, böbrek tübülleri, çizgili kaslarda ve hayvanların uçma kaslarında, oksidatif fosforilasyon olaylarının arttığı durumlarda çok sık dizilmiş kristalara rastlanır.En fazla krista sayısı böceklerin uçma kaslarında bulunur.
Mitokondriler labil yapılı organellerdir. Farklı etmenlerin etkisinde değişikliğe uğrarlar. Zarar gören hücrelerde parçalanarak küçük taneler haline geçerler. İçlerinde yoğun maddeler birikerek daha şişkin hale gelebilirler. Değişiklikler bir sınırı aşınca mitokondriler dejenerasyona uğrarlar. Bundan sonra meydana gelen değişiklikler artık reversibl değildir. Reversibl olmayan değişiklikler üç şekilde gelişir:
1) Ya küçük tanecikler halinde parçalanarak dispersiyon oluştururlar
2) Veya şişerek geniş vakuoller haline gelirler
3) Ya da mitokondrilerin hiyalin granülleri haline dönüşmesiyle şişmiş, geniş, kitlesel birikintiler meydana getirirler.
Böylelikle çoğunlukla hücre ölür. Normal hücrelerde bir tür lisosom (sitolisosom) oluşumuyla mitokondriler dejenerasyona uğrayara k otolize olurlar. Mitokondrilerin bir başka hali de bir araya gelip birleşerek kondriosfer denen daha büyük cisimleri meydana getirmeleridir. Hacim ve şekil değiştirmeleri, kimyasal maddeler ve osmatik basınç değişiklikleriyle sağlanabilir.